Wersja z 2018-08-04
List do komórki. Hormony zwierzęce
![]()
![]()
![]()
Wstęp
Hormony to substancje aktywne biologicznie. Wytwarzane są one przez komórki gruczołowe i wydzielane do krwi; kontaktują się z receptorami komórek docelowych i działają na ich procesy życiowe.
W oparciu o wybrane cechy wyróżnia się odpowiednie grupy hormonów.
Wśród hormonów występują substancje drobnocząsteczkowe – przede wszystkim pochodne aminokwasów i steroidy – oraz wielkocząsteczkowe hormony peptydowe.
Hormony (w ujęciu klasycznym) to substancje powstające w gruczołach dokrewnych, a substancje powstające w komórkach rozproszonych określa się mianem hormonów tkankowych.
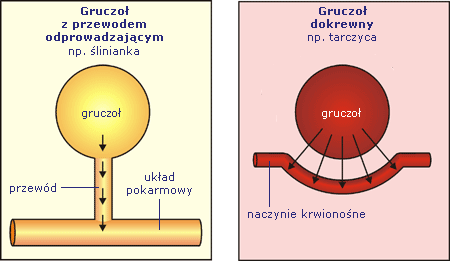
Dwa typy gruczołów
Pod względem zasięgu działania hormony dzieli się na:
Hormony wpływające na wydzielanie innych hormonów to hormony sterujące, czyli tropowe. Odgrywają one szczególną rolę w regulacji hormonalnej, budują osie sprzężeń. Hormony, których wydzielanie od nich zależy, zwrotnie wpływają na wydzielanie tychże hormonów sterujących, regulując ich stężenie we krwi.
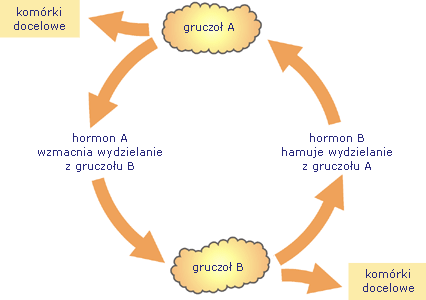
Zasada sprzężenia zwrotnego w wydzielaniu hormonów
Wymienione cechy pozwalają układowi hormonalnemu kontrolować pracę narządów, przyczyniając się do utrzymania równowagi procesów zachodzących w organizmie przy jednoczesnej zdolności do adaptacji do zmieniających się warunków życia.
Jak hormon działa na komórkę?
Hormon przenosi informację między komórkami. Komórki, które odbierają daną informację posiadają odpowiedni aparat receptorowy dla danego hormonu. Połączenie hormonu z receptorem wywołuje sekwencję zdarzeń, czyli właściwą reakcję komórki. Uczestniczą w tym procesie informatory komórkowe podporządkowane hormonom, dlatego określa się te substancje nazwą informatory wtórne lub przekaźniki drugiego rzędu. Substancje indukujące ich działanie – hormony: peptydowe, neuropeptydy, klasyczne neuroprzekaźniki, hormony steroidowe, cytokiny, eikozanoidy i cząsteczki adhezji międzykomórkowej – to informatory pierwotne (przekaźniki I rzędu).
Każdy rodzaj hormonu ma w komórkach docelowych właściwy receptor, z którym może się połączyć. To połączenie wywołuje reakcję komórki docelowej. Szczegóły budowy hormonu i jego receptora umożliwiają przeniesienie informacji w organiźmie – można to porównać do listu w zaadresowanej kopercie, która trafia do adresata mieszkającego w określonym miejscu. Podobnie, jak osoby które mają adres inny, niż zapisany na kopercie nie otrzymają listu, tak komórki nie posiadające receptora dla danego hormonu, nie będą reagowały na jego wpływ.
Hormony steroidowe łatwo przechodzą przez zbudowaną z substancji lipidowych (tłuszczowców) błonę komórkową, więc mogą połączyć się z receptorami zlokalizowanymi w cytoplazmie. Białkowe hormony mają cząsteczki o dużych rozmiarach. Nie przechodzą więc przez błonę komórek – dlatego receptory dla tych informatorów wbudowane są w błonę komórkową.
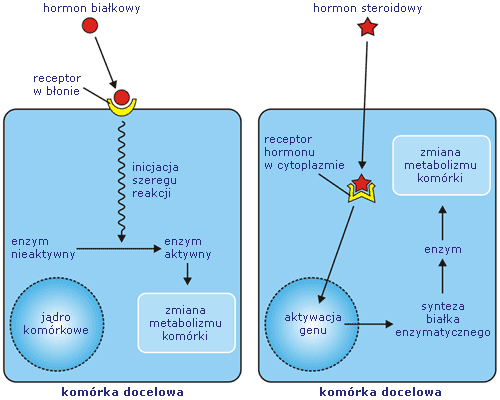
Sposób w jaki komórka przyjmuje „list” w postaci hormonu steroidowego i białkowego (peptydowego)
Komórka docelowa odpowiada aktywacją genów, co wpływa na syntezę białek (enzymów), lub inicjuje kaskadę reakcji wpływających na zmianę aktywności enzymów, które odpowiadają za poszczególne chemiczne procesy życiowe komórki.
Literatura dla dociekliwych:
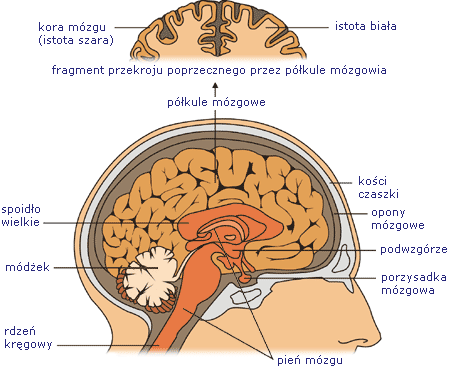
Mózg. Jaki jest stosunek topograficzny przysadki mózgowej do szkieletu?
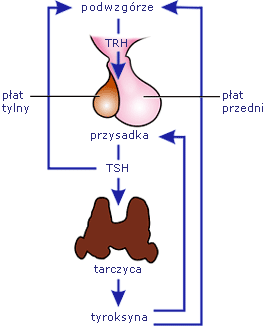
Podwzgórze i przysadka.
W jaki sposób podwzgórze komunikuje się z przysadką?
Hormony jąder wielkokomórkowych uwalniane w płacie tylnym przysadki mózgowej
Hormony hamujące uwalnianie (statyny)
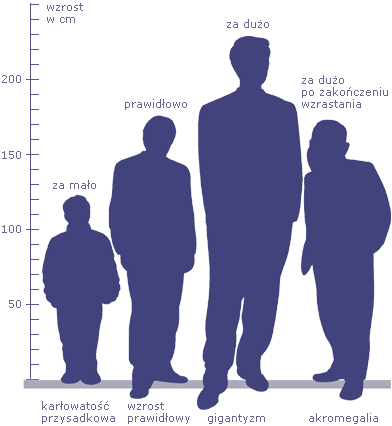
Gigantyzm i karłowatość. Nadmiar hormonu wzrostu pojawiający się przed zakończeniem okresu wzrostu organizmu prowadzi do gigantyzmu, niedobór zaś – do karłowatości. Natomiast nadmiar tego hormonu po okresie wzrostu skutkuje akromegalią. Z którą sytuacją zatem wiąże się nadmierny wzrost ciała?
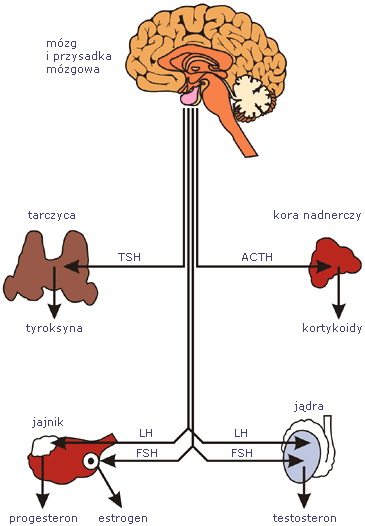
Oś podwzgórze-przysadka-tarczyca.
Jakie hormony uczestniczą we wzajemnej regulacji wydzielania tyroksyny?
ACTH jest polipeptydem złożonym z 39 aminokwasów, pochodzącym z proopiomelanokortyny (POMC) zbudowanej z 285 aminokwasów. Działa na receptory błonowe komórek warstwy pasmowatej kory nadnerczy. Interakcja hormonu z receptorem pobudza układ cyklaza adenylanowa-cAMP. Pod wpływem ACTH wzrasta zawartość enzymów i mRNA w komórkach kory nadnerczy, co jest przejawem troficznego charakteru wpływu ACTH na te komórki.
ACTH hamuje wydzielanie CRH, jest także czynnikiem stymulującym dla melanocytów skóry.
Płat przedni przysadki mózgowej
Hormony części pośredniej
Hormony płata tylnego
Hormony tylnego płata przysadki mózgowej to oksytocyna i wazopresyna opisane w podrozdziale „Hormony jąder wielkokomórkowych uwalniane w płacie tylnym przysadki mózgowej” opisującym hormony podwzgórzowe. (Część 3 artykułu – kliknij TUTAJ).
IV. HORMONY NADNERCZY
Hormony rdzenia nadnerczy:
Hormony kory nadnerczy
Hormony kory nadnerczy produkowane są w komórkach kory nadnerczy (bogatych w witaminę C) pod wpływem zwiększenia stężenia cAMP indukowanego przez ACTH. Cholesterol ulega przekształceniu w pregnenolon – substancję wyjściowa dla szeregu reakcji syntez hormonów steroidowych – w reakcji katalizowanej przez 20,22-desmolazę cholesterolową.
Warstwa kłębkowata kory gruczołów nadnerczowych wydziela głównie mineralokortykoidy, warstwa pasmowata – glikokortykoidy, siatkowata – androgeny.
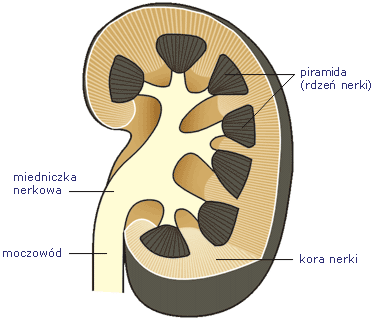
VI. HORMONY TARCZYCY
Hormony pęcherzyków tarczycowych
Hormon komórek przypęcherzykowych c
VII. HORMON GRUCZOŁÓW PRZYTARCZYCZNYCH
VIII. HORMONALNA POSTAĆ WITAMINY D3
Hormonalna postać witaminy D3 sprzyja mineralizacji i odnowie kości, utrzymuje stałe stężenie wapnia we krwi.
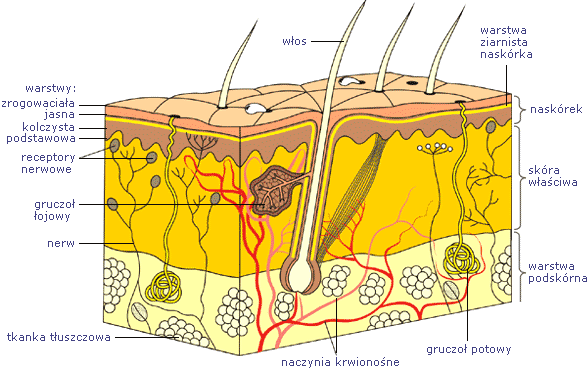
Skóra. Która część ciała jest najbardziej eksponowana na światło?
Cholekalcyferol jest wytwarzany w skórze ssaków pod wpływem promieni nadfioletowych światła słonecznego z 7-dehydrocholesterolu. Witamina D3 jest następnie transportowana ze swoistą globuliną wiążącą witaminę D (DBP) – składnik osocza krwi – do wątroby, gdzie w reticulum endoplazmatycznym gładkim hepatocytów pod wpływem 25-hydroksylazy ulega zamianie w prohormon 25-hydroksycholekalcyferol 25(OH)D3. Ten ostatni jest transportowany przez DBP do kanalików proksymalnych nerek i tam ulega zamianie w 1,25(OH)2D3 lub 24,25(OH)2D3. Pierwszy z wymienionych, powstający pod wpływem parahormonu lub utraty fosforanów z ustroju, bardziej aktywnie przyczynia się do nasilenia wchłaniania wapnia z przewodu pokarmowego. Z tego powodu jest uznawany za właściwy hormon.
1,25(OH)2D3 dyfunduje z nerek i łączy się z DBP.
Witamina D3 poprzez receptory wewnątrzkomórkowe wzmaga m.in. syntezę mRNA i DBP cytozolowego w komórkach nabłonka jelita. Dzięki temu enterocyty wchłaniają wapń bardziej wydajnie.
Witamina D3 ułatwia działanie PTH na kości.
IX. HORMONY WYSP TRZUSTKI
Hormony wysp trzustki to glukagon, insulina, hormon hamujący uwalnianie hormonu wzrostu (SRIF), polipeptyd trzustkowy (PP).
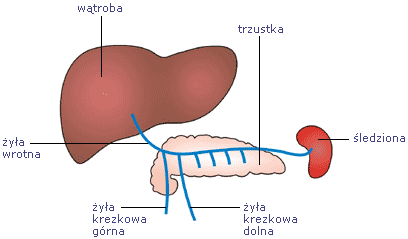
Dorzecze żyły wrotnej wątroby.
Jakie znaczenie ma przepływ krwi z jelit i trzustki do wątroby?
Hormony jajników
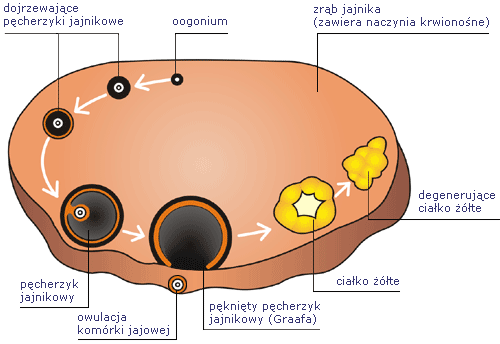
Jajnik. Kiedy powstaje i gdzie się znajduje ciałko żółte?
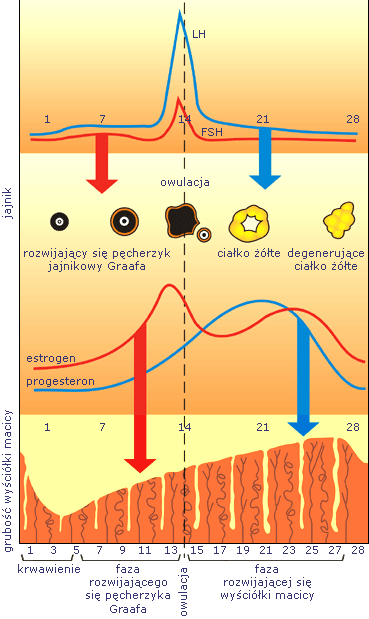
Cykl miesiączkowy. Jaki jest udział hormonów przysadki w cyklu miesiączkowym?
Hormony łożyska
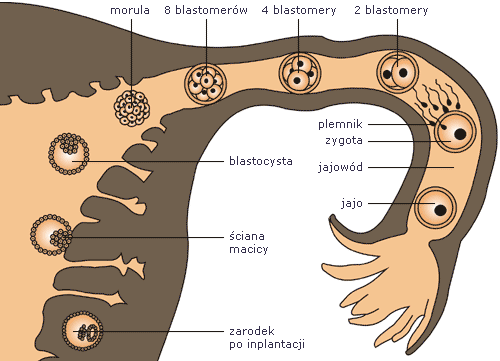
Rozwój zarodka od zapłodnienia do implantacji w ścianie macicy
hPL składa się z 191 aminokwasów i wykazuje podobieństwo do hormonu wzrostu i prolaktyny.
Jest wytwarzany od piątego tygodnia ciąży. W niewielkim stopniu wpływa na utrzymanie ciałka żółtego i podobnie jak hormon wzrostu wykazuje działanie anaboliczne polegające na nasilaniu syntezy białek i uwalnianiu wolnych kwasów tłuszczowych. Dzięki temu zużycie glukozy zmniejsza się. hPL jest czynnikiem tropowym dla gruczołów mlecznych i podobnie jak prolaktyna, lecz w mniejszym stopniu, wzmaga wytwarzanie mleka.
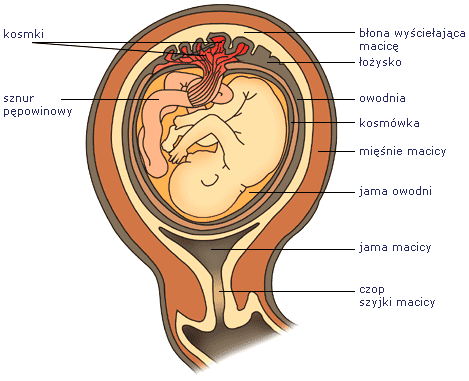
Łożysko. Jakie znaczenie ma szczególna lokalizacja łożyska?
Progesteron powstaje z cholesterolu w szeregu reakcji, w którym występuje produkt pośredni – pregnenolon. Progesteron w około 2/3 przechodzi do krwiobiegu matki. Pozostała część (wyłączając niewielki udział progesteronu rozkładanego w łożysku) trafia do krwi płodu.
Przechodzenie progesteronu z łożyska do macicy umożliwia wpływ tego hormonu na utrzymanie ciąży.
Progesteron jest dla płodu prekursorem kortyzolu, kortyzonu i aldosteronu, które powstają w korze nadnerczy płodu.
NEUROPRZEKAŹNIKI
Neuroprzekaźniki przenoszą informacje między komórkami nerwowymi. Uwalniane do synaps przez komórki przedsynaptyczne wzbudzają potencjały czynnościowe komórek odbierających sygnały.
Hormony regulujące metamorfozę u owadów holometabolicznych
![]()
![]()
![]()